The lack of agreement of researchers cannot be denied with regard to the Date and Place of Origin of Haplotype J1. However, the usual suspect origin is from the Levant (Lebenon, Israel, Jordan, and eastern Egypt) with two dispersion events toward North Africa and the Arabian Plate, regardless who does the research.
The J or J-12f2k Haplogroup is the grandparent of J1 and has an estimated age is 31.7±12.8 Kyr, or 31,700 years old with possible range of possible ages from 18,900 years to 44,500 years old. Of course, the more recent paper (Karafet et al, 2008, pg.7 of 8) with demography-independent age estimates puts the IJ, parent of J, with the age of 38,500 years(Range:30,500–46,200).That means you have to clip off that J claim to account for the lower ranges of IJ.
The estimated age of the J1/M267 subclade's age of origin setting began with "24.1±9.4 Kyr" which translates from researcher jargon to 24,100 years before present, plus or minus 9,400 years. That means the J1 age range of 14,700 to 33,500 overlaps both of the parent ranges of J and IJ. That is an inconsistancy.
In this study dating J1 to 14,700 to 33,500 years old, only included 93 individuals classified as J-12f2k that were not J2. The most common ancestor of the J-M267 branch 93 chromosomes out of a population of 257 males,typed 275 men from five populations in Algeria, Tunisia, and Egypt with a set of 119 binary markers and 15 microsatellites from the Y chromosome. They state, "...we have analyzed the results together with published data from Moroccan populations found in both the Middle East and North Africa (and including our J* chromosomes).
To put that into perspective, 24,100 years ago was the last height when ice sheets were at their maximum extension. While humans were using tools, their teeth show they still used them as a tool and a spare hand in the Mesolithic Natufian culture from 12,500 to 9,500 BC in the Levant.
To compare with 9,400 years, about 9,400 years ago was long before there was domestication of plants and animals. The land bridge between the British Isles was just beginning to fill in and there was no written language. The oldest written language is Sumerian from about 5,000 years ago (5.0 Kybp), so every event before that is "prehistoric."
The parent clade of J or M304, which is offspring of the J Haplogroup and parent of J1 and J1, is estimated to be 20,000 years before present, so, the 24.1 Kybp date of origin for the J1-M267 given lacks some verifiable authority. It is better set at 18,200 years before present(C. Cinniog˘ lu et al, 2003, Pg.131) or 18,000 years(Di Giacomo et al,2004, Table 3., Pg 364).
Another part of the problem is the data gives information in generations rather than years, but the researchers put years instead of generations in their papers. Some researchers us 25 years per generation and others 30 for their assumed time between generations. Some genealogists use 20 years as an assumed time for a generation. The time between generations has the factor of when the first and last children were born affecting the average outcomes. Those factors have been in flux over the last 9,000 years and have drastically changed in the last 400 years. The difference is similar to the difference between the old English (pound) versus the new metric (kilogram) systems.
J1-M267 Y lineage marks climate-driven pre-historical human displacements
European Journal of Human Genetics (2009) 17, 1520–1524; doi:10.1038/ejhg.2009.58; published online 15 April 2009 http://www.nature.com/ejhg/journal/v17/n11/full/ejhg200958a.html Sergio Tofanelli1,14, Gianmarco Ferri2,14, Kazima Bulayeva3, Laura Caciagli1, Valerio Onofri4, Luca Taglioli1, Oleg Bulayev3, Ilaria Boschi5, Milena Alù2, Andrea Berti6, Cesare Rapone6, Giovanni Beduschi2, Donata Luiselli7, Alicia M Cadenas8, Khalid Dafaallah Awadelkarim9,10, Renato Mariani-Costantini10, Nasr Eldin Elwali9, Fabio Verginelli10, Elena Pilli11, Rene J Herrera8, Leonor Gusmão12, Giorgio Paoli1 and Cristian Capelli13
Calculations under the coalescent model for J1 haplotypes bearing the Cohanim motif gave time estimates that place the origin of this genealogy around 6.2 Kybp (95% CI: 4.5–8.6 Kybp), earlier than previously thought,4 and well before the origin of Judaism (David Kingdom, ~2.0 Kybp).This misinformation in the paper calls into question the bias of the authors for this paper.
NOTE: 2.0 Kybp or 2,000 years ago would be during the Roman occupation of Israel, long after the rein of David.Grabbe 2008, pp. 225–6. puts Judah into 8th-7th century BC. The Cohen lineage, however, begins with Aaron, the brother of Moses.The Jewish calendar's epoch (reference date), 1 Tishrei 1 AM, is equivalent to Monday, 7 October 3761 BCE (or 5.771 Kybp) in the proleptic Julian calendar, the equivalent tabular date (same daylight period) and is about one year before the traditional Jewish date of Creation on 25 Elul AM 1, based upon the Seder Olam Rabbah of Rabbi Yossi ben Halafta, a 2nd century CE sage.The Jewish year 5771 AM began on 9 September 2010 (1 Tishrei or Rosh Hashanah) and, since it is a 13-month year, will end on 28 September 2011 (29 Elul).The Book of Kings relates how a "law of Moses" was discovered in the Temple during the reign of king Josiah (r. 641–609 BC)William Dever agrees with the Canaanite origin of the Israelites but allows for the possibility of some immigrants from Egypt among the early hilltop settlers, leaving open the possibility of a Moses-like figure in Transjordan ca 1250-1200 BCE or 3250 to 3200 or 3.250 Kybp. New papers on molecular biology add the J-P58 or J1c3 subclade requirement to the Cohen Modal Haplotype, which may make it younger. Ultimately, it is unlikely Aaron was the founder of his haplotype. That might be pushed back further to Abraham.
However, a wide range of times since the most recent common ancestor (TMRCAs) has been proposed for J1 and its subclades (between 36 and 10 KyBP), and different conflicting scenarios have been depicted to explain their current distribution.3, 5, 6, 7, 8, 9
Semino et Al 2004, Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe Hum Genet (2004) 115: 357–371 DOI 10.1007/s00439-004-1168-9 http://www.familytreedna.com/pdf/HaploJ.pdfPg.358:We also present here, for the first time, a novel method for comparative dating of lineages, free of assumptions of STR mutation rates,Table 3,pg.364:1 YMRCA=8,400+/-12,600 SD,3 BATWING a(subhaplogroups treated separately)=18,000(7,590–33,990)a.Age in years rounded to the nearest hundred, assuming 30 years/generation (Zerjal et al. 2002, 2003)
Arredi B, Poloni ES, Paracchini S et al: A predominantly Neolithic origin for Y-chromosomal DNA variation in North Africa. Am J Hum Genet 2004; 75: 338–345. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1216069/Pg.342:An origin for haplogroup J in the Middle East has been proposed (Semino et al. 2004 and references therein); the TMRCA of the J-M267 branch, found in both the Middle East and North Africa (and including our J* chromosomes), was estimated at 24.1 ± 9.4 KY and must predate its spread. This is consistent with our 95% TMRCA estimate of 4.4–11.1 KY for the North African chromosomes.
Cadenas AM, Zhivotovsky LA, Cavalli-Sforza LL, Underhill PA, Herrera RJ: Y-chromosome diversity characterizes the Gulf of Oman. Eur J Hum Genet 2008; 16: 374–386. http://www.nature.com/ejhg/journal/v16/n3/full/5201934a.htmlhttp://www.nature.com/ejhg/journal/v16/n3/pdf/5201934a.pdfPg.376:Haplogroup-specific expansion times were estimated for select binary haplogroups (J1-M267, R1a1-M198, E3b1a-M78 and E3b1c-M123) by the linear expansion method. This procedure assumes a stepwise mutation model 45and a mean STR mutation rate of 0.00069 per STR locus per generation 46 with a 25-year intergeneration time as performed in previous studies. 4,12,47Pg.382:Based on binary and STR markers, the greatest degree of differentiation for J1-M267 is detected in the Levant with two distinct demographic dispersals generating its current distribution. A higher observed STR diversity of this clade among Europeans and Ethiopians in comparison to populations of North Africa points to its arrival to Ethiopia and Europe during Neolithic times with a more recent appearance in the latter.5858 Semino O, Magri C, Benuzzi G et al: Origin, diffusion, and differentiation of Y-chromosome haplogroups E and J: inferences on the neolithization of Europe and later migratory events in the Mediterranean area. Am J Hum Genet 2004; 74: 1023 – 103, Pg.383:Median BATWING expansion times based on Y-STR data for the Omani (2.3 ky; 95% CI: 0.6– 29.2) J1-M267 chromosomes 4 indicate a more recent arrival to the South Arabian populations as compared to the older expansion times obtained for the Egyptian (6.4 ky; 95% CI: 0.6–278.5) 4 and Turkish (15.4 ky; 95% CI: 0.4– 604.8)12representatives of this haplogroup. Conversely, in the present study, Y-STR age estimates based on the method described by Zhivotovsky et al 46 generated much older values for the J1-M267 haplogroup in Yemen, Qatar and UAE (9.772.4, 7.472.3 and 6.471.4 ky, respectively) than seen in the Omani,4 consistent with an earlier arrival to the region during the Neolithic.
Chiaroni J, King RJ, Underhill P: Correlation of annual precipitation with human Y chromosome diversity and the emergence of Neolithic agriculture and pastoral economies in the fertile crescent. Antiquity 2008; 82: 281–289. Access Restricted.
Saudi Arabian Y-Chromosome diversity and its relationship with nearby regions Khaled K Abu-Amero1 , Ali Hellani2 , Ana M González3 , Jose M Larruga3 , Vicente M Cabrera3 and Peter A Underhill4 http://www.biomedcentral.com/1471-2156/10/59
Page 6 of 9:
The peculiar distribution of J2-M67 in Arabia could be explained assuming maritime contacts from classical Mediterranean cultures. The presence in Saudi Arabia of three males harbouring underived J1-M304 (Both J1 and J2)chromosomes is intriguing. It could be that they came together with the J1-M267 or J2-M172 expansive waves, or they could represent the remnants of an old and geographically widespread Palaeolithic substrate. This type of underived chromosomes has been detected rarely in Turkey [21], in Oman and in the eastern Mediterranean area [34]. However, as the critical Levantine region has
not yet been adequately dissected for J1, it seems premature to favor any of these hypotheses. The geographic pattern and most probable origin of the Y-chromosome haplogroup J in Arabia faithfully mirrors those found for the most prevalent J and R0a mtDNA haplogroups in the same region [7-9,12]. In addition, J1-M267 divergence age calculated for Saudi Arabia (11.6 ± 2 kya) and Yemen (11.3 ± 2 kya) are also very coincidental with those calculated for J1b (11.1 ± 8.4 kya) and R0a1 (9.6 ± 2.9 kya) in
Saudi Arabia [7,8]. It is worth mentioning that J1-M267 ages in Saudi Arabia and Yemen are significantly older than those obtained for UAE and Qatar (Table 3)[15] and for Oman [14] pointing to a terrestrial more than to a maritime colonization. It has been suggested that Yemen could be a center of expansion for mtDNA haplogroup R0a [9].
Table 3: Y-haplogroup J1-M267 variance and divergence times deduced from Y-STR loci
Population sample size k17 a k14 b M-267 variance T(ky) Divergence time (ky) Mean ± SE
UAE c 57 40 33 0.16 5.81 6.81 ± 1.53
Qatar c 42 33 28 0.19 6.71 7.27 ± 1.83
Yemen c 45 41 40 0.27 9.69 11.27 ± 2.03
Saudi Arabia 48 41 39 0.29 10.37 11.59 ± 1.93
Arabian Peninsula 192 149 129 0.24 8.85 10.78 ± 1.65
a Haplotype number using 17 Y-STR loci
b Haplotype number using 14 Y-STR loci
c Estimated from the Cadenas et al. (2007) data
18,200 Years Before Present
Excavating Y-chromosome haplotype strata in Anatolia Hum Genet,volume 114 2004
Pg.131:Table 2 Y-chromosome haplogroup variance and expansion times based on ten STR loci:J-304J-M304 n=175 0.56 20.0 T (kyr) ; J1-M267,n=47, Variance=0.51, T (kyr)=18.6 Pg.133Haplogroup J and the transition to agriculture Although the entire J-M304 (J1+J2) clade demonstrates a large microsatellite variance that under a continuous growth model dates to round 20 kyr, consistent with the LGM, the BATWING exponential growth model reveals a more recent post-LGM expansion (13.9 kyr). This secondary expansion originates from a low effective population size (n=184) and may indicate that the J clade in Turkey began to participate in demographic expansions during the onset of sedentism in Anatolia and the Levant; e.g., the Natufians (Bar-Yosef 1998). Previously, J clade representatives would have been accumulating STR diversity via genetic drift within various small groups of mobile hunter-gathers during the LGM.http://ychrom.invint.net/upload/iblock/129/Cinnioglu%202003%20Excavating%20Y-chromosome%20haplotype%20strata%20in%20Anatolia.pdf doi=10.1007/s00439-003-1031-4 | pages=127 | pmid=14586639 | first1=Cengiz | last2=King | first2=Roy | last3=Kivisild | first3=Toomas | last4=Kalfoglu | first4=Ersi | last5=Atasoy | first5=Sevil | last6=Cavalleri | first6=Gianpiero L. | last7=Lillie | first7=Anita S. | last8=Roseman | first8=Charles C. | last9=Lin | first9=Alice A. | issue=2}}
[1]
Evidence of Bias, Lack of Accuracy, External and Internal Validity in J1 Dating from "J1-M267 Y lineage marks climate-driven pre-historical human displacements"[edit]
J1-M267 Y lineage marks climate-driven pre-historical human displacements
"However, a wide range of times since the most recent common ancestor (TMRCAs) has been proposed for J1 and its subclades (between 36 and 10 KyBP), and different conflicting scenarios have been depicted to explain their current distribution.3, 5, 6, 7, 8, 9", http://www.nature.com/ejhg/journal/v17/n11/full/ejhg200958a.html
3 Semino O, Magri C, Benuzzi G et al: Origin, diffusion and differentiation of Y-chromosome haplogroups E and J: inferences on the neolithization of Europe and later migratory events in the Mediterranean area. Am J Hum Genet 2004; 74: 1023–1034.
Figure 2. Phylogeny and frequency distributions of Hg J and its main subclades (panels A–F). The numbering of mutations is according to the YCC (YCC 2002; Jobling and Tyler-Smith 2003). To the left of the phylogeny, the ages (in 1,000 years) of the boxed mutations are reported, with their SEs (Zhivotovsky et al. 2004). With the exception of the age relative to the 12f2 mutation, which has been estimated as TD (with V0=0) between the combined data of the two sister clades Hg J-M267 and Hg J-M172, the other values have been determined as ASD, as described in figure 1. (Figure 1 depicts a phylogenetic tree indicating: J-12f2=31.7+/-12.8 ky; 267=24.1+/-9.4 ky; 172=18.5+/-3.5; et cetera) http://www.sciencedirect.com/science/article/pii/S0002929707643663#fig1
5 Di Giacomo F, Luca F, Popa LO et al: Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe. Hum Genet 2004; 115: 357–371.
Table 3 Estimates of age of haplogroup J, J sub-haplogroups, and mutation rate at five STR loci, obtained with three independent methods(st. dev. standard deviation, NA not applicable),Estimation method,p12f2, M267:1.YMRCAa=8,400, 2.st. dev.b=12,600, 3.BATWINGa(subhaplogroups treated separately)=18,000, 4.5th–95th centileb= 7,590–33,990, 5.BATWINGa(subhaplogroups and UEPs)=8,300–11,300, 5th–95th centileb=4,260–23,070,aAge in years rounded to the nearest hundred, assuming 30 years/generation (Zerjal et al. 2002, 2003),bAge as above, rounded to the nearest tenthhttp://www.familytreedna.com/pdf/HaploJ.pdf
6 Arredi B, Poloni ES, Paracchini S et al: A predominantly Neolithic origin for Y-chromosomal DNA variation in North Africa. Am J Hum Genet 2004; 75: 342.
(Semino et al. 2004 and references therein); the TMRCA of the J-M267 branch, found in both the Middle East and North Africa (and including our J* chromosomes), was estimated at 24.1+/-9.4 KY and must predate its spread. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1216069/pdf/AJHGv75p338.pdf
Table 3 Y-Haplogroup variance, expansion and coalescence times based on Y-microsatellite loci,Divergence timea(ky)b, Qatar-J1-M267=7.4+/-2.3, UAE-J1-M267=6.4+/-1.4, Yemen-J1-M267=9.7+/-2.4,aDivergence time based on SNP-STR coalescence method (Zhivotovsky46,51).,bBased on 15 Y-STR loci http://www.nature.com/ejhg/journal/v16/n3/pdf/5201934a.pdf
8 Zalloua PA, Xue Y, Khalife J et al: Y-chromosomal diversity in Lebanon is structured by recent historical events. Am J Hum Genet 2008; 82: 873–882.**http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2427286/pdf/main.pdf Article regards distribution rather than ages.
9 Chiaroni J, King RJ, Underhill P: Correlation of annual precipitation with human Y chromosome diversity and the emergence of Neolithic agriculture and pastoral economies in the fertile crescent. Antiquity 2008; 82: 281–289.
Geographical Structure of the Y-chromosomal Genetic Landscape of the Levant: A coastal-inland contrast FROM: John Scharf TO: Dr. Pierre Zalloua The Lebanese American University[edit]
It is not clear why you chose to cite and base your paper on Tofanelli et al. He starts by fogging the date of origin of the origin of J1/M267 by stating a range of 36,000 to 10,000 ybp while citing papers that do not back the upper end of the range being beyond 33,990. Further, the lower range is pushed down to as low as 4,260 by the papers he cites.
He also says Calculations under the coalescent model for J1 haplotypes bearing the Cohanim motif gave time estimates that place the origin of this genealogy around 6.2 Kybp (95% CI: 4.5–8.6 Kybp), earlier than previously thought,4 and well before the origin of Judaism (David Kingdom, ~2.0 Kybp). That he makes this departure for religious reasons and makes this glaring historic error I go into detail at the bottom of my notes.
The claim of J1 coming from Africa has issues as well. In "Y-chromosome diversity characterizes the Gulf of Oman. Eur J Hum Genet 2008; 16: 374–386." the paper states the age of the J1 in Turkey to be (15.4 ky; 95% CI: 0.4– 604.8). This article also points out dates of origin for M267 in Yemen, Qatar and UAE (9.772.4, 7.472.3 and 6.471.4 ky, respectively). Are we to understand that J1/M267 jumps over the Levant and lands in Anatolia before going to the Arabian Plate?
My letter to It is not clear why you chose to cite and base your paper on Tofanelli et al. He starts by fogging the date of origin of the origin of J1/M267 by stating a range of 36,000 to 10,000 ybp while citing papers that do not back the upper end of the range being beyond 33,990. Further, the lower range is pushed down to as low as 4,260 by the papers he cites.
He also says Calculations under the coalescent model for J1 haplotypes bearing the Cohanim motif gave time estimates that place the origin of this genealogy around 6.2 Kybp (95% CI: 4.5–8.6 Kybp), earlier than previously thought,4 and well before the origin of Judaism (David Kingdom, ~2.0 Kybp). That he makes this departure for religious reasons and makes this glaring historic error I go into detail at the bottom of my notes.
The claim of J1 coming from Africa has issues as well. In "Y-chromosome diversity characterizes the Gulf of Oman. Eur J Hum Genet 2008; 16: 374–386." the paper states the age of the J1 in Turkey to be (15.4 ky; 95% CI: 0.4– 604.8). This article also points out dates of origin for M267 in Yemen, Qatar and UAE (9.772.4, 7.472.3 and 6.471.4 ky, respectively). Are we to understand that J1/M267 jumps over the Levant and lands in Anatolia before going to the Arabian Plate?
NOTE: The notes reference are listed on this page.
Listed here are notable ethnic groups by Y-DNAhaplogroups based on relevant studies. The data is presented in two columns for each haplogroup with the first being the sample size (n) and the second the percentage in the haplogroup designated by the column header. The samples are taken from individuals identified with the ethnic and linguistic designations in the first two columns.
This user page needs to be updated. Please help update this user page to reflect recent events or newly available information. Relevant discussion may be found on the talk page. (February 2021)
^Van Oven M, Van Geystelen A, Kayser M, Decorte R, Larmuseau HD (2014). "Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome". Human Mutation. 35 (2): 187–91. doi:10.1002/humu.22468. PMID24166809. S2CID23291764.
^K-M2313*, which as yet has no phylogenetic name, has been documented in two living individuals, who have ethnic ties to India and South East Asia. In addition, K-Y28299, which appears to be a primary branch of K-M2313, has been found in three living individuals from India. See: Poznik op. cit.; YFull YTree v5.08, 2017, "K-M2335", and; PhyloTree, 2017, "Details of the Y-SNP markers included in the minimal Y tree" (Access date of these pages: 9 December 2017)
^ Haplogroup S, as of 2017, is also known as K2b1a. (Previously the name Haplogroup S was assigned to K2b1a4.)
^ Haplogroup M, as of 2017, is also known as K2b1b. (Previously the name Haplogroup M was assigned to K2b1d.)
^ abcdefghijklmnopqrstuvwxyzaaabacCadenas, Alicia M; Zhivotovsky, Lev A; Cavalli-Sforza, Luca L; Underhill, PA; Herrera, RJ (2008). "Y-chromosome diversity characterizes the Gulf of Oman". European Journal of Human Genetics. 16 (3): 374–86. doi:10.1038/sj.ejhg.5201934. PMID17928816.
^ abcdefghiU.-D. Immela, M. Erhumab, T. Mustafac, M. Kleibera and M. Klintschara, "Population genetic analysis in a Libyan population using the PowerPlex 16 system," International Congress Series Volume 1288, April 2006, Pages 421-423
^ abcdMichael E. Weale, Levon Yepiskoposyan, Rolf F. Jager, Nelli Hovhannisyan, Armine Khudoyan, Oliver Burbage-Hall, Neil Bradman, Mark G. Thomas, "Armenian Y chromosome haplotypes reveal strong regional structure within a single ethno-national group," Human Genetics (2001) 109 : 659–674.
^ abcdefgDoron M. Behar, Daniel Garrigan, Matthew E. Kaplan et al., "Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations," Human Genetics (2004) 114 : 354–365.
^Lobov. Y chromosome analysis in subpopulations of Bashkirs from Russia
^ abcdefV. N. Kharkov, V. A. Stepanov, S. P. Feshchenko, S. A. Borinskaya, N. K. Yankovsky, and V. P. Puzyrev, "Frequencies of Y Chromosome Binary Haplogroups in Belarussians," Russian Journal of Genetics, Vol. 41, No. 8, 2005, pp. 928–931.
^ abcdefQamar Raheel; et al. "Y-Chromosomal DNA Variation in Pakistan". American Journal of Human Genetics. 70 (1107–1124): 2002. {{cite journal}}: Unknown parameter |author-separator= ignored (help)
^ abcdefghijklmnopqrst[1] Estimating Scandinavian and Gaelic Ancestry in the Male Settlers of Iceland - Agnar Helgason et al., 2000, Am. J. Hum. Genet. 67:697–717, 2000
^ abcdefF. Luca, F. Di Giacomo, T. Benincasa et al., "Y-Chromosomal Variation in the Czech Republic," American Journal of Physical Anthropology 132:132–139 (2007).
^ abcdefghijklmnopqrstuElizabeth T Wood, Daryn A Stover, Christopher Ehret et al., "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes," European Journal of Human Genetics (2005) 13, 867–876. cf. "Appendix A: Y Chromosome Haplotype Frequencies."
^ abcdefLaitinen Virpi, Lahermo Päivi, Sistonen Pertti, Marja- , Savontaus Liisa (2002). "Y-Chromosomal Diversity Suggests that Baltic Males Share Common Finno-Ugric-Speaking Forefathers". Human Heredity. 53 (2): 68–78. doi:10.1159/000057985. PMID12037406.{{cite journal}}: CS1 maint: multiple names: authors list (link)
^ abcdefghij[2] Genetic evidence for different male and female roles during cultural transitions in the British Isles - James F. Wilson et al., 5078–5083, PNAS, April 24, 2001, vol. 98, no. 9
^ abcde[3] Y Chromosome Evidence for Anglo-Saxon Mass Migration - Michael E. Weale et al.,Mol. Biol. Evol. 2002;19:1008–1021.
^ abcdefghijklmnopKarafet, Tatiana; Xu, Liping; Du, Ruofu; et al. "Paternal Population History of East Asia: Sources, Patterns, and Microevolutionary Processes". American Journal of Human Genetics. 69 (615–628): 2001. {{cite journal}}: Explicit use of et al. in: |first3= (help)
^ abcdefghijklmYali Xue, Tatiana Zerjal, Weidong Bao, Suling Zhu, Qunfang Shu, Jiujin Xu, Ruofu Du, Songbin ***, Pu Li, Matthew E. Hurles, Huanming Yang, Chris Tyler-Smith, "Male demography in East Asia: a north-south contrast in human population expansion times," Genetics 2006.
^ abcdefghMichael F. Hammer, Tatiana M. Karafet, Hwayong Park, Keiichi Omoto, Shinji Harihara, Mark Stoneking and Satoshi Horai, "Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes," Journal of Human Genetics Volume 51, Number 1 / January, 2006.
^ abcdefghijklmnopqrstuvwxyzaaOleg Balanovsky, Siiri Rootsi, Andrey Pshenichnov et al., "Two Sources of the Russian Patrilineal Heritage in Their Eurasian Context," American Journal of Human Genetics 82, 236–250, January 2008.
^ abcdefghijKarlsson, Andreas O; Wallerström, Thomas; Götherström, Anders; Holmlund, Gunilla (2006). "Y-chromosome diversity in Sweden – A long-time perspective". European Journal of Human Genetics. 14 (8): 963–70. doi:10.1038/sj.ejhg.5201651. PMID16724001.
^ abcdefghijklmnopqrSanchez, Juan J; Hallenberg, Charlotte; Børsting, Claus; Hernandez, Alexis; Morling, Niels (2005). "High frequencies of Y chromosome lineages characterized by E3b1, DYS19-11, DYS392-12 in Somali males". European Journal of Human Genetics. 13 (7): 856–66. doi:10.1038/sj.ejhg.5201390. PMID15756297.
^ abcdefT. Lappalainen, V. Laitinen, E. Salmela et al., "Migration Waves to the Baltic Sea Region," Annals of Human Genetics (2008).
^ abcZerjal, Tatiana; Wells, R. Spencer; Yuldasheva, Nadira; Ruzibakiev, Ruslan; Tyler-Smith, Chris. "A Genetic Landscape Reshaped by Recent Events: Y-Chromosomal Insights into Central Asia". American Journal of Human Genetics. 71 (466–482): 2002.
^ abcdeRuixia Zhou, Daqun Yang, Hua Zhang, Weiping Yu, Lizhe An, Xilong Wang, Hong Li, Jiujin Xu, and Xiaodong Xie, "Origin and evolution of two Yugur sub-clans in Northwest China: a case study in paternal genetic landscape," Annals of Human Biology (2008), 35:2, 198 — 211.
The Aurignacian culture is an archaeological culture of the Upper Palaeolithic, located in Europe and southwest Asia. It lasted broadly within the period from ca. 45,000 to 35,000 years ago in terms of conventional radiocarbon dating, or between ca. 47,000 and 41,000 years ago in terms of the most recent calibration of the radiocarbon timescale. [1] The name originates from the type site of Aurignac in the Haute Garonne area of France. The most critical single discovery is that of the so called Egbert skeleton from Ksar Akil 10 km northeast of Beirut in Lebanon, embedded in deposits overlain by Levantine Aurignacian industries.(Ziffer 1978 harvnb error: no target: CITEREFZiffer1978 (help))This is a fully modern human in both cranial and postcranial terms, between 40,000 and 45,000 years old. [2]
The Antelian culture is an Upper Paleolithic phase of the Levant (Syria, Lebanon, Palestine) that evolves from Emirian. The most important innovation in this period is the incorporation of some typical elements of Aurignacian, like some types of burins and narrow blade points that resemble the European type of Font-Yves. [3]
The Kebaran or Kebarian culture was an archaeological culture in the eastern Mediterranean area (c. 18,000 to 10,000 BC), named after its type site, Kebara Cave south of Haifa. The Kebaran were a highly mobile nomadic population, composed of hunters and gatherers in the Levant and Sinai areas who utilized microlithic tools.The Kebaran is the last Upper Paleolithic phase of the Levant (Syria, Lebanon, Palestine). The Kebarans were characterized by small, geometric microliths, and were thought to lack the specialized grinders and pounders found in later Near Eastern cultures. The Kebaran is preceded by the Athlitian phase of the Antelian and followed by the proto-agrarian Natufian culture of the Mesolithic. The Kebaran is also characterised by the earliest collecting of wild cereals, known due to the uncovering of grain grinding tools. It was the first step towards the Neolithic Revolution. The Kebaran people are believed to have practice dispersal to upland environments in the summer, and aggregation in caves and rockshelters near lowland lakes in the winter. This diversity of environments may be the reason for the variety of tools found in their toolkits. Situated in the Terminal Pleistocene, the Kebaran is classified as an Epipalaeolithic society. They are generally thought to have been ancestral to the later Natufian culture that occupied much of the same range.[4]
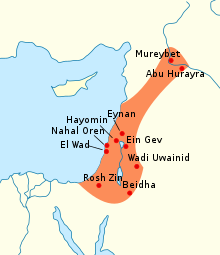
The spread of Natufian cultureThe Natufian culture was a Mesolithic culture that existed from 12,500 to 9,500 BC in the Levant, a region in the Eastern Mediterranean. It was unusual in that it was sedentary, or semi-sedentary, before the introduction of agriculture.The Natufian communities are possibly the ancestors of the builders of the first Neolithic settlements of the region, which may have been the earliest in the world.[5] There is some evidence for the deliberate cultivation of cereals, specifically rye, by the Natufian culture, at the Tell Abu Hureyra site, the site for earliest evidence of agriculture in the world.[6] Generally, though, Natufians made use of wild cereals. [7] Animals hunted include gazelles.[8]The term "Natufian" was coined by Dorothy Garrod who studied the Shuqba cave in Wadi an-Natuf, Israel, about halfway between Tel Aviv and Ramallah.Radiocarbon dating places this culture from the terminal Pleistocene to the very beginning of the Holocene, from 12,500 to 9,500 BC.[9][10]The period is commonly split into two subperiods: Early Natufian (12,500–10,800 BC) and Late Natufian (10,800–9500 BC). The Late Natufian most likely occurred in tandem with the Younger Dryas (10,800 to 9500 BC). In the Levant, there are more than a hundred kinds of cereals, fruits, nuts and other edible parts of plants, and the flora of the Levant during the Natufian period was not the dry, barren, and thorny landscape of today, but parkland and woodland.[11]
The '''Pre-Pottery Neolithic B''' Pre-Pottery Neolithic B (PPNB) is a division of the Neolithic developed by Dame Kathleen Kenyon during her archaeological excavations at Jericho in the southern Levant region.Sites from this period found in the Levant utilizing rectangular floor plans and plastered floor techniques were found at Ain Ghazal, Yiftahel (western Galilee), and Abu Hureyra (Upper Euphrates).[12] The period is dated to between ca. 10700 and ca. 8000 BP or 8700 - 6000 BCE.
The Khiamian (also referred to as El Khiam or El-Khiam) is a period of the Near-Eastern Neolithic, marking the transition between the Natufian and the Pre-Pottery Neolithic A. It dates from about 10,000 to 9,500 BCE.[13]
Pre-Pottery Neolithic A (PPNA for short) denotes the first stage in early Levantine Neolithic culture, dating around 9500 to 8500 BC[14]. Archaelogical remains are located in the Levantine and upper Mesopotamian region of the Fertile Crescent. The culture is characterized by small circular mud brick dwellings, the cultivation of crops, the hunting of wild game, and unique burial customs in which bodies were buried below the floors of dwellings.[15] The Pre-Pottery Neolithic A and the following Pre-Pottery Neolithic B were originally defined by Kathleen Kenyon in the type site of Jericho (Palestine). During this time, pottery was yet unknown. They precede the ceramic Neolithic (Yarmukian). PPNA succeeds the Natufian culture of the Epipaleolithic (Mesolithic).
Yarmukian pottery vessel, Sha'ar HaGolan.The Yarmukian Culture is a Neolithicculture of the ancient Levant. It was the first culture in Prehistoric Israel and one of the oldest in the Levant to make use of pottery. The Yarmukian derives its name from the Yarmouk River which flows near its type site at Sha'ar HaGolan, a kibbutz at the foot of the Golan Heights. The first Yarmukian settlement was unearthed at Megiddo during the 1930s, but was not identified as a distinct Neolithic culture at the time. At Sha'ar HaGolan, in 1949, Prof. Moshe Stekelis first identified the Yarmukian Culture, a Pottery Neolithic culture that inhabited parts of Israel and Jordan.[16] The site, dated to ca. 6400–6000 BC (calibrated), is located in the central Jordan Valley, on the northern bank of the Yarmouk River. Its size is circa 20 hectares, making it one of the largest settlements in the world at that time. Although other Yarmukian sites have been identified since, Sha'ar HaGolan is the largest, probably indicating its role as the Yarmukian center.[17]
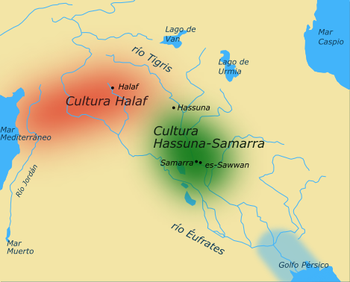
The geographic location of the Chalcolithic Halaf culture in relation to the contemporaneous Hassuna culture.Halaf culture, is a prehistoric culture which developed from Neolithic III at Tell Halaf without any strong break. The Tell Halaf site flourished from about 6100 to 5400 BCE, a period of time that is referred to as the Halaf period. The Halaf culture was succeeded in northern Mesopotamia by the Ubaid culture. The site was then abandoned for a long period.Dryland farming was practiced by the population. This type of farming was based on exploiting natural rainfall without the help of irrigation, in a similar practice to that still practiced today by the Hopi people of Arizona. Emmer wheat, two-rowed barley and flax were grown. They kept cattle, sheep and goats.
^P.Mellars, Archeology and the Dispersal of Modern Humans in Europe: Deconstructing the Aurignacian, Evolutionary Anthropology, vol. 15 (2006), pp. 167–182.
^Settlement Patterns and Intrasite Variability in the Central Negev, Israel†
Anthony E. Marks
Article first published online: 28 OCT 2009
DOI: 10.1525/aa.1971.73.5.02a00210
1971 American Anthropological Association
http://onlinelibrary.wiley.com/doi/10.1525/aa.1971.73.5.02a00210/pdf
^Mellaart, James (1976), "Neolithic of the Near East" (MacMillan)
^Barker G (2002) Transitions to farming and pastoralism in North Africa, in Bellwood P, Renfrew C (2002), Examining the Farming/Language Dispersal Hypothesis, pp 151–161.
^Moore, Andrew M. T. (2000). Village on the Euphrates: From Foraging to Farming at Abu Hureyra. Oxford: Oxford University Press. ISBN019510806X. {{cite book}}: Unknown parameter |coauthors= ignored (|author= suggested) (help)
^Bar-Yosef O (1987) Pleistocene connections between Africa and SouthWest Asia: an archaeological perspective. The African Archaeological Review; Chapter 5, pg 29-38
^Cite error: The named reference Kottak2005 was invoked but never defined (see the help page).
^Cite error: The named reference Munro2003 was invoked but never defined (see the help page).
^Ofer Bar-Yosef, The Natufian culture and the Early Neolithic: Social and economic trends in Southwestern Asia, chapter 10 in Peter Bellwood and Colin Renfrew (eds.), Examining the Farming/Language Dispersal Hypothesis (2002), p.114.
^Cite error: The named reference Bar-Yosef_159 was invoked but never defined (see the help page).
^Amihai Mazar, Archaeology of the Land of the Bible:10,000 - 586 BCE, Doubleday: New York, 1992, 45.
^C. Calvet. 2007. Zivilisationen – wie die Kultur nach Sumer kam. Munich. p. 126.
^Cite error: The named reference PNAS09 was invoked but never defined (see the help page).
^Mithen, Steven (2006). After the ice : a global human history, 20,000-5000 BC (1st Harvard University Press pbk. ed. ed.). Cambridge, Mass.: Harvard University Press. p. 63. ISBN0674019997. {{cite book}}: |edition= has extra text (help)